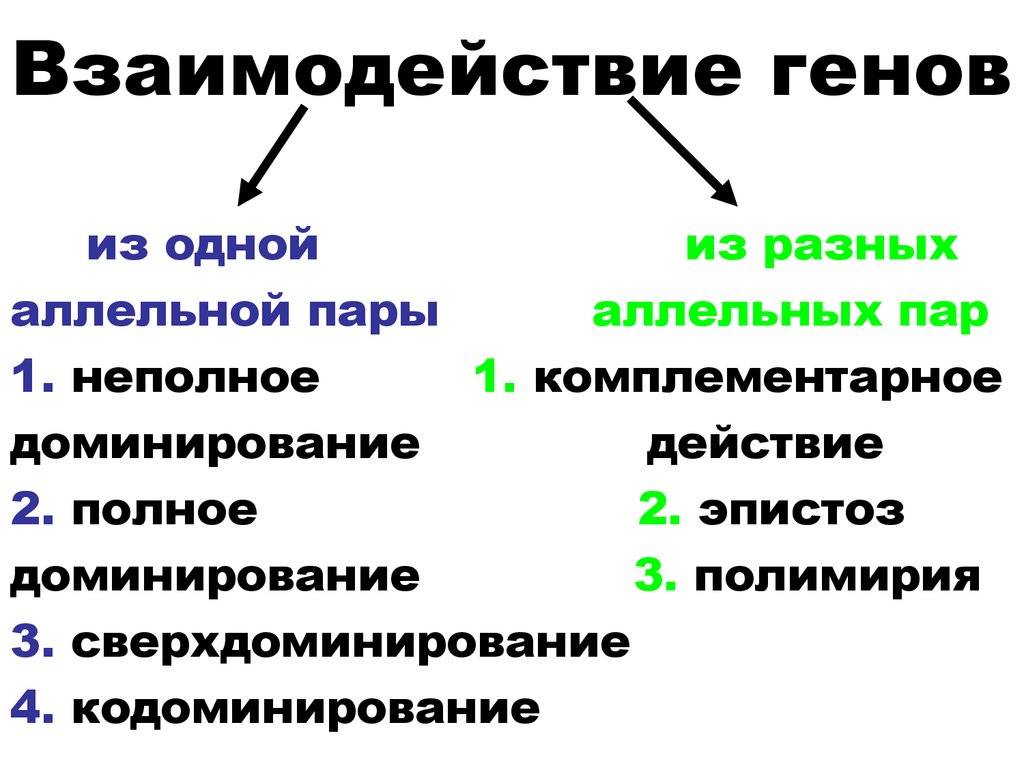
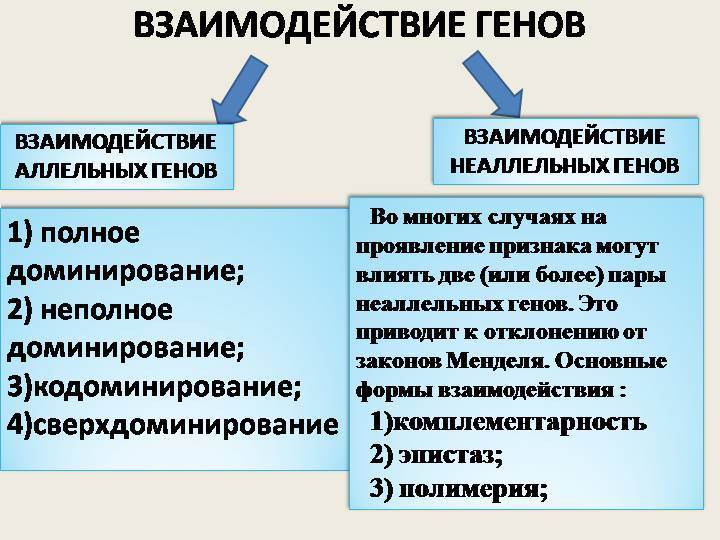
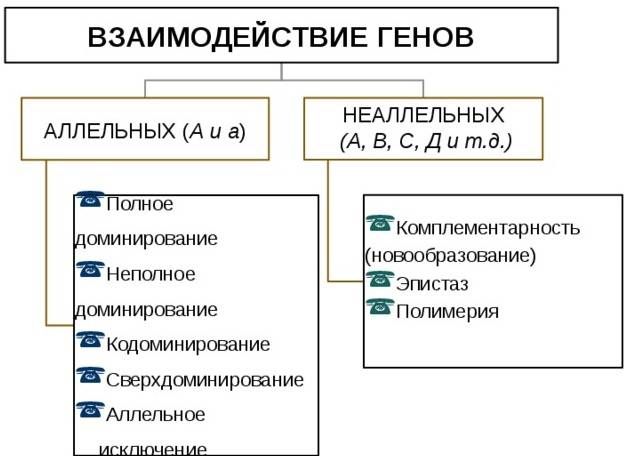
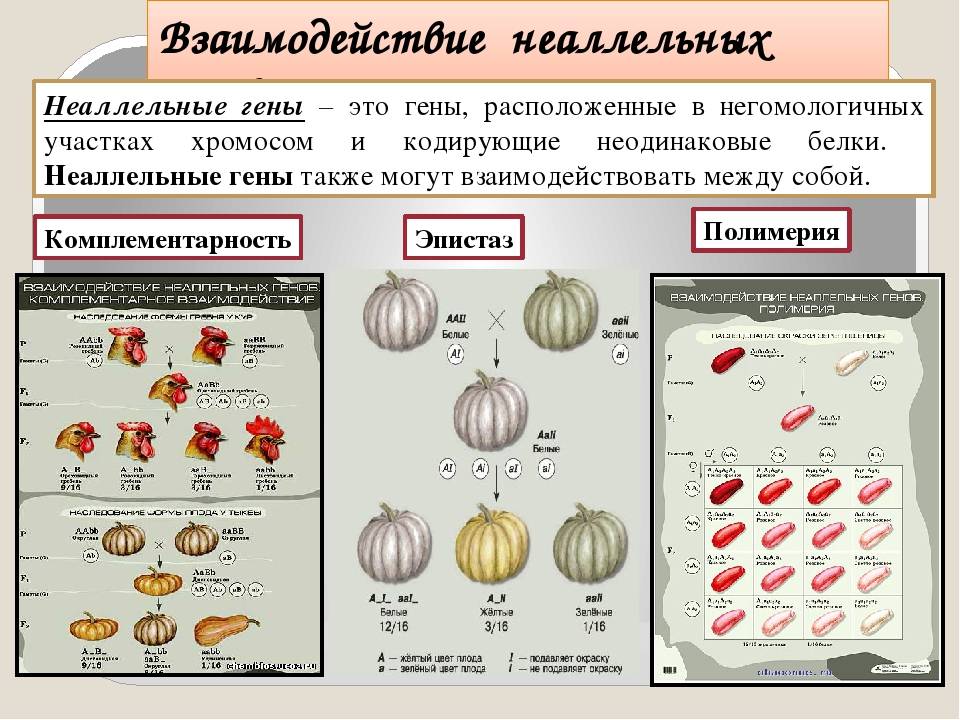
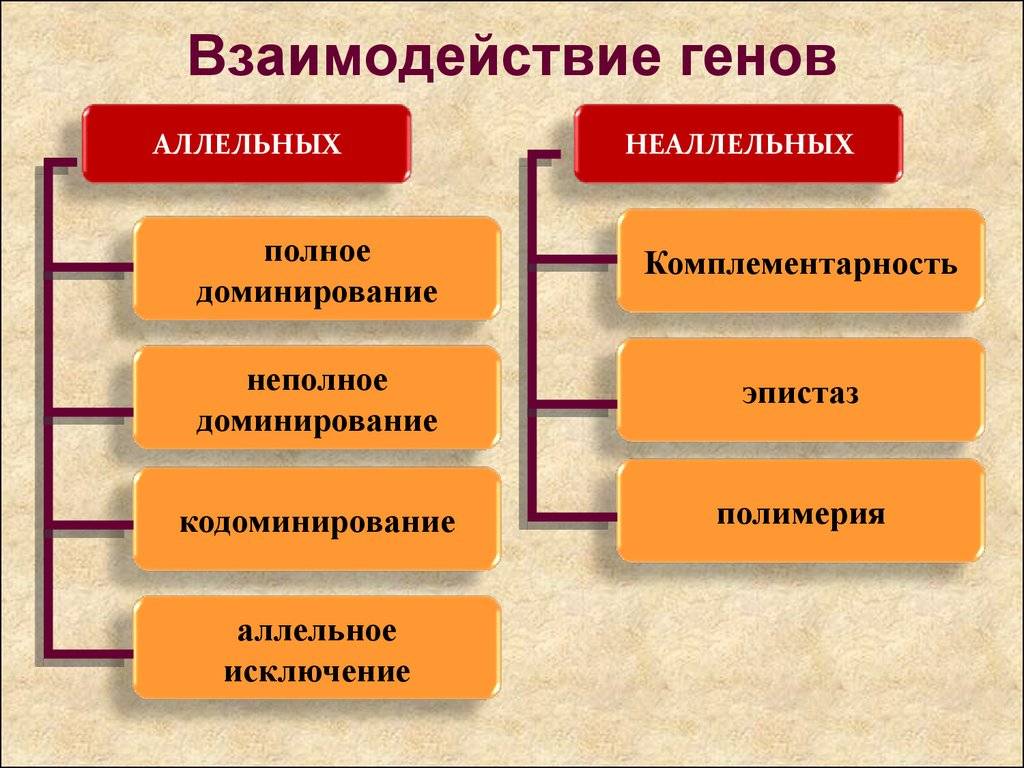
Взаимодействие неаллельных генов
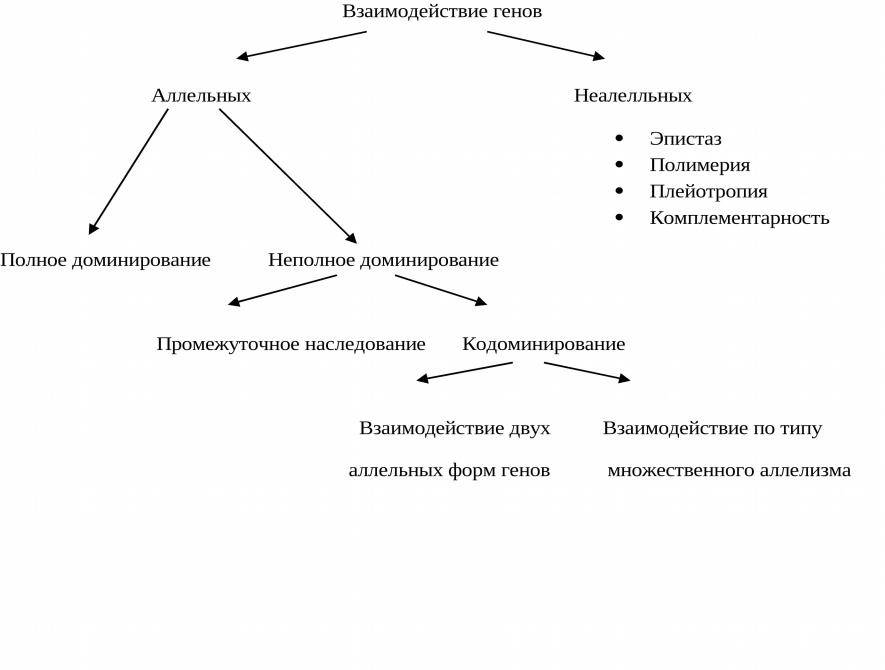
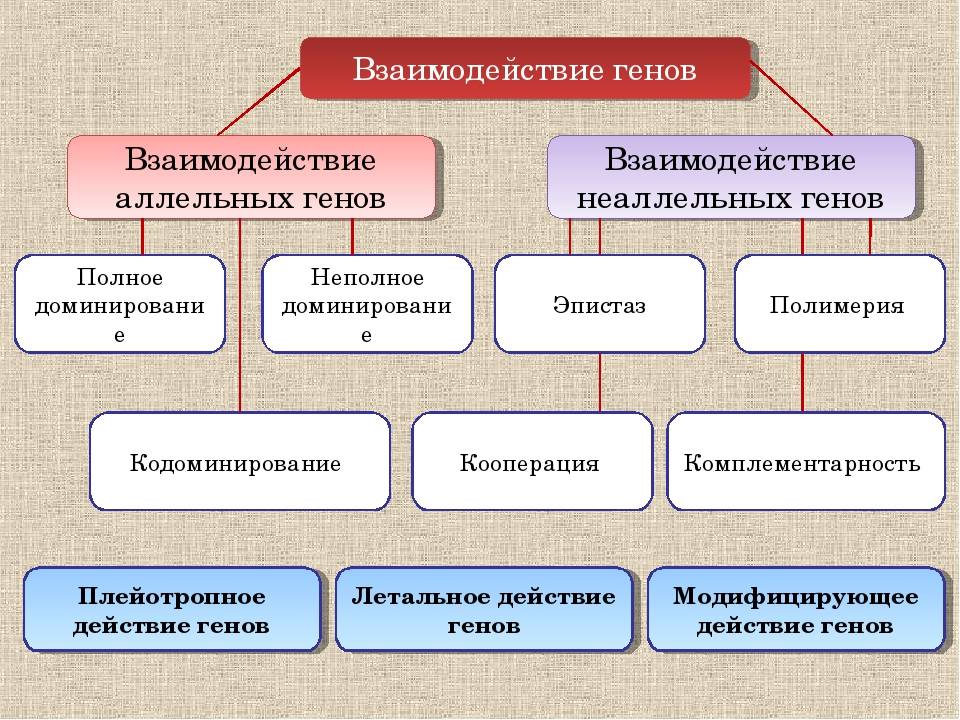
Неаллельное (или межаллельное) взаимодействие — это взаимодействие аллелей разных генов, т.е. генов, располагающихся в негомологичных хромосомах или разных локусах гомологичных хромосом.
■ Неаллельное взаимодействие генов приводит к модификации менделевского расщепления по фенотипу 9 : 3 : 1, т.е. к появлению в потомстве гетерозиготы иных расщеплений, например 9 : 3 : 4; 9 : 6 : 1; 12 : 3 : 1 и др.
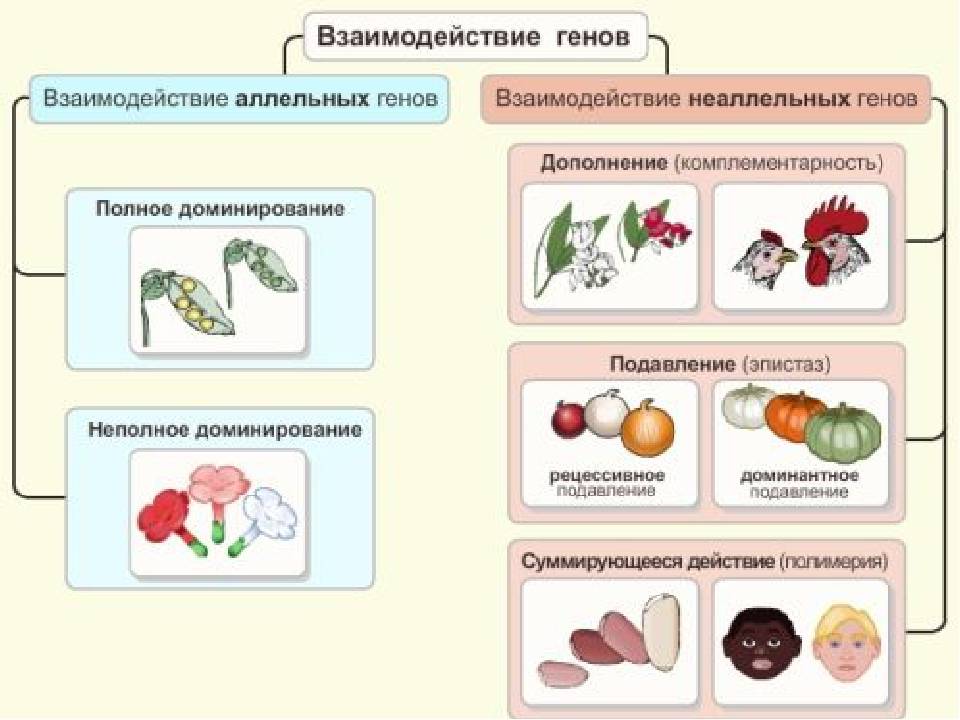
❖ Основные типы межаллельных взаимодействий:
■ комплементарность;
■ эпистаз;
■ полимерия.
Замечание: комплементарное и эпистатическое взаимодействия возникают в тех случаях, когда признак контролируется одной парой неаллельных генов.
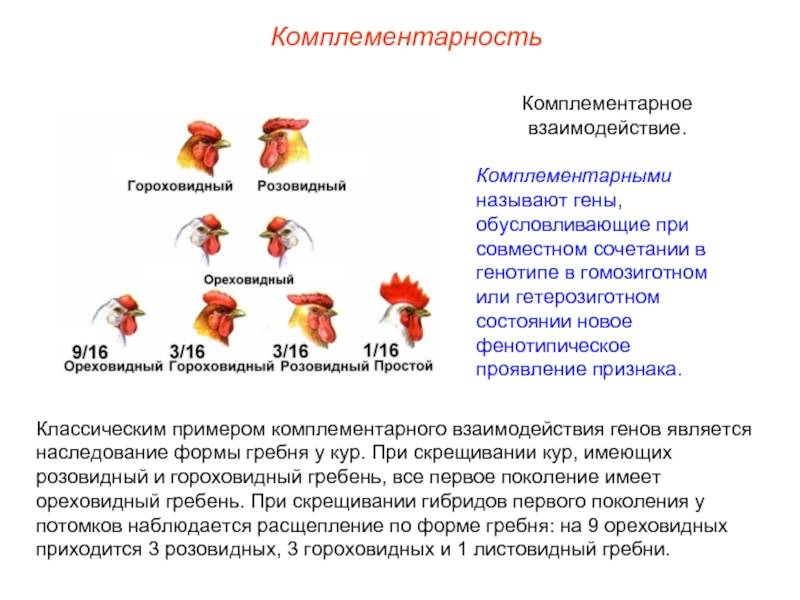
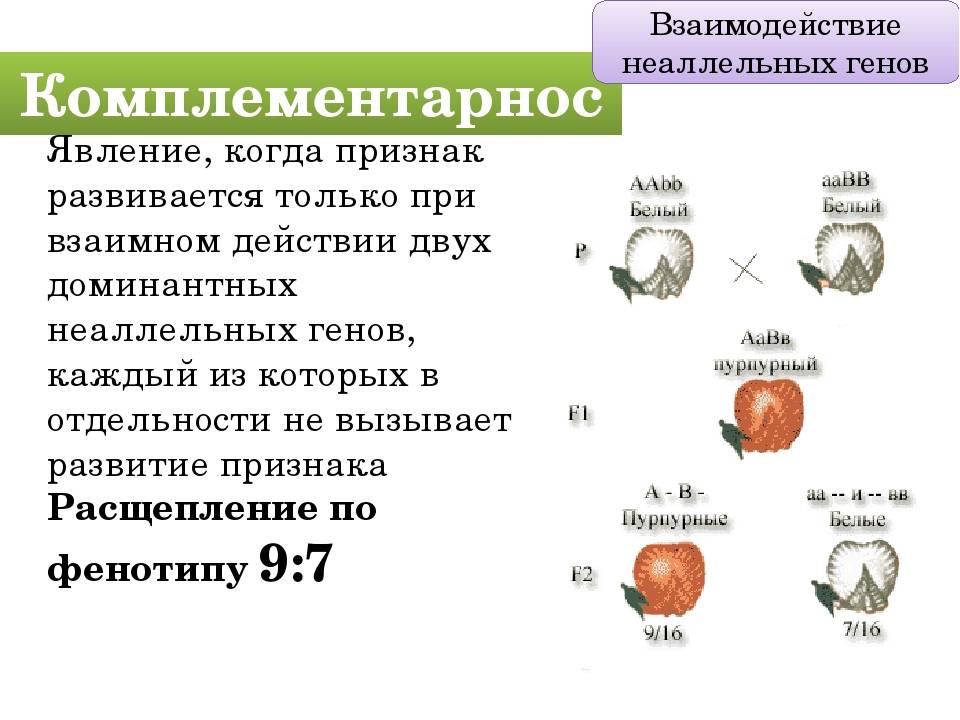
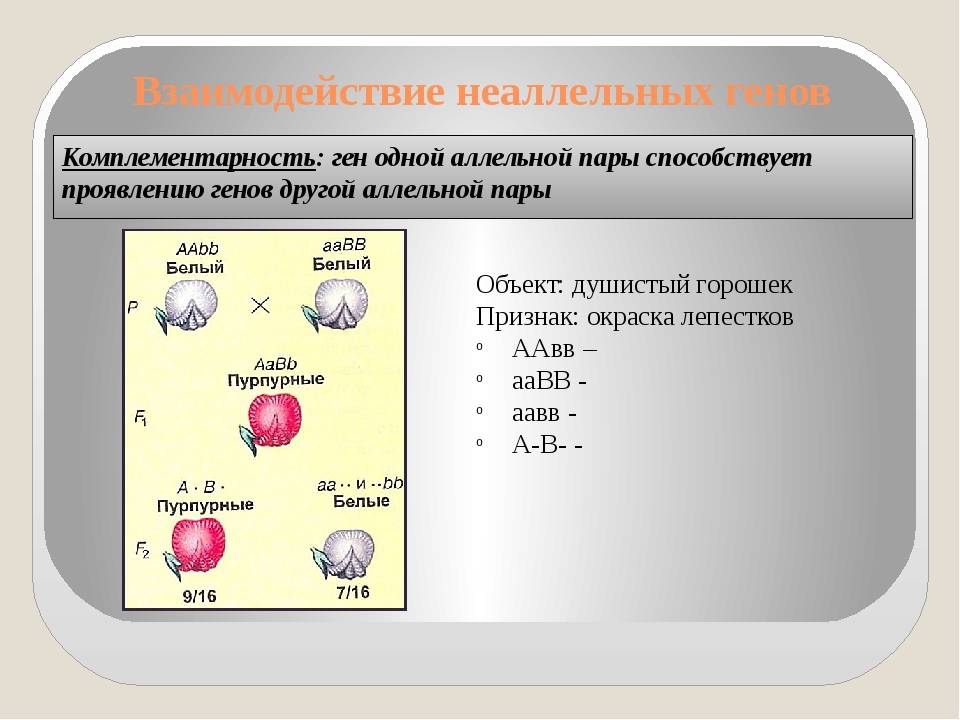
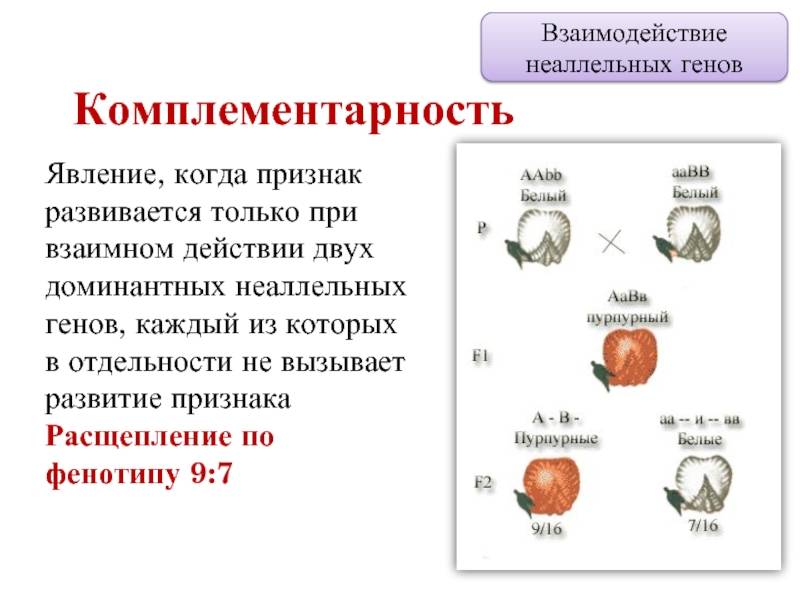
Комплементарное, или дополнительное, взаимодействие — такой тип межаллельного взаимодействия генов, при котором одновременное присутствие в генотипе гибрида доминантных генов разных аллельных пар приводит к появлению нового признака, отсутствующего у обоих родителей.
Пример: наследование окраски цветков душистого горошка (родительские растения с генотипами А-вв, ааВ- имеют белые цветки, гибриды с генотипом А-В- пурпурные; см. таблицу).
Замечание: знак «-» в формуле генотипа означает, что это место может занимать как доминантный, так и рецессивный аллель.
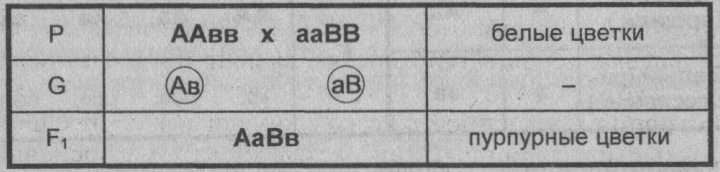
Объяснение: пурпурный пигмент образуется с помощью специального фермента, который синтезируется только при наличии обоих доминантных генов: как А, так и В. Цветки родительских особей имеют белый цвет, так как в генотипе каждого из них присутствует только один из этих генов.
Во втором поколении при самоопылении, обеспечивающем равновероятное (случайное) образование гамет и зигот разного типа, наблюдается расщепление по фенотипу в соотношении пурпурной и белой окраски цветков как 9 : 7 (9 пурпурных: A-В- и 7 белых: ЗА-Bb, ЗааВ-. 1aabb).
Эпистаз — такой тип межаллельного взаимодействия генов, при котором аллели одного гена подавляют проявление аллельной пары другого гена, и подавляемый признак не проявляется.
Супрессор (или ген-ингибитор) — ген, подавляющий действие других неаллельных генов. Супрессором может быть как доминантный, так и рецессивный ген.
Доминантный эпистаз — эпистаз, в котором супрессором является доминантный ген. При доминантном эпистазе во втором поколении наблюдается расщепление фенотипа 12:3:1 или 13:3.
Рецессивный эпистаз (криптомерия) — эпистаз, в котором супрессором является рецессивный ген. При рецессивном эпистазе во втором поколении наблюдается расщепление по фенотипу 9:3:4.
Пример эпистаза: наследование окраски шерсти у домашних кроликов. Синтез черного пигмента детерминирует рецессивный ген с, доминантная аллель I другого гена является супрессором, подавляя действие гена с. Тогда кролики с генотипами C-I-, ccl-будут белыми, кролики с генотипами C-ii — серыми, а с генотипом ссii — черными.
Многие признаки контролируются двумя и более парами неаллельных генов (называемых в этом случае полимерными).
Полимерия — взаимодействие нескольких неаллельных полимерных генов. При полимерии степень выраженности фенотипического признака часто зависит от числа полимерных генов, ответственных за его проявление. При кумулятивной полимерии действие генов суммируется; примеры: масса тела, молочность крупного рогатого скота, яйценоскость кур, некоторые параметры умственных способностей человека и др. При некумулятивной полимерии степень проявления признака от числа доминантных генов в генотипе не зависит (пример: оперенность ног у кур).
Плейотропия — зависимость нескольких признаков от одного гена. Каждый плейотропный ген оказывает какое-то основное действие, но модифицирует проявление других генов.
Полимерия
Это вид взаимодействия двух и более пар неаллельных генов, доминантные аллели которых однозначно влияют на развитие одного и того же признака. Полимерное действие генов может быть кумулятивным и некумулятивным. При кумулятивной полимерии интенсивность значения признака зависит от суммирующего действия генов: чем больше доминантных аллелей, тем больше степень выраженности признака. При некумулятивной полимерии количество доминантных аллелей на степень выраженности признака не влияет, и признак проявляется при наличии хотя бы одного из доминантных аллелей. Полимерные гены обозначаются одной буквой, аллели одного локуса имеют одинаковый цифровой индекс, например А1а1А2а2А3а3.
Кумулятивная полимерия имеет место при наследовании окраски зерновок пшеницы, чешуек семян овса, роста и цвета кожи человека и т.д.
| P | A1A1A2A2черные чешуйки | × | a1a1a2a2белые чешуйки |
| Типы гамет | A1A2 | a1a2 | |
| F1 | A1a1A2a2серые чешуйки, 100% | ||
| P | A1a1A2a2серые чешуйки | × | A1a1A2a2серые чешуйки |
| Типы гамет | A1A2 A1a2 a1A2 a1a2 | A1A2 A1a2 a1A2 a1a2 |
| A1A2 | A1a2 | a1A2 | a1a2 | ||
| A1A2 | A1A1A2A2черные | A1A1A1a2темно-серые | A1a1A2A2темно-серые | A1a1A2a2серые | |
| A1a2 | A1A1A2a2темно-серые | A1A1a2a2серые | A1a1A2a2серые | A1a1а2a2светло-серые | |
| a1A2 | A1a1A2A2темно-серые | A1a1A2a2серые | a1a1A2A2серые | a1a1A2a2светло-серые | |
| a1a2 | A1a1A2a2серые | A1a1a2a2светло-серые | a1a1A2a2светло-серые | a1a1a2a2желтые |
Черные чешуйки семян у овса — 1/16, темно-серые чешуйки семян у овса — 4/16, серые чешуйки семян у овса — 6/16, светло-серые чешуйки семян у овса — 4/16, желтые чешуйки семян у овса — 1/16. Расщепление по фенотипу 1:4:6:4:1.
Некумулятивная полимерия имеет место при наследовании формы плодов пастушьей сумки.
| P | A1A1A2A2треугольные | × | a1a1a2a2овальные |
| Типы гамет | A1A2 | a1a2 | |
| F1 | A1a1A2a2треугольные, 100% | ||
| P | A1a1A2a2треугольные | × | A1a1A2a2треугольные |
| Типы гамет | A1A2 A1a2 a1A2 a1a2 | A1A2 A1a2 a1A2 a1a2 |
| A1A2 | A1a2 | a1A2 | a1a2 | ||
| A1A2 | A1A1A2A2треугольные | A1A1A1a2треугольные | A1a1A2A2треугольные | A1a1A2a2треугольные | |
| A1a2 | A1A1A2a2треугольные | A1A1a2a2треугольные | A1a1A2a2треугольные | A1a1а2a2треугольные | |
| a1A2 | A1a1A2A2треугольные | A1a1A2a2треугольные | a1a1A2A2треугольные | a1a1A2a2треугольные | |
| a1a2 | A1a1A2a2треугольные | A1a1a2a2треугольные | a1a1A2a2треугольные | a1a1a2a2овальные |
Треугольная форма плодов у пастушьей сумки — 15/16, овальная форма плодов у пастушьей сумки — 1/16.
Плейотропия — множественное действие генов. Плейотропное действие генов имеет биохимическую природу: один белок-фермент, образующийся под контролем одного гена, определяет не только развитие данного признака, но и воздействует на вторичные реакции биосинтеза других признаков и свойств, вызывая их изменение.





Плейотропное действие генов впервые было обнаружено Г. Менделем, который установил, что у растений с пурпурными цветками всегда имелись красные пятна в пазухах листьев, а семенная кожура была серого или бурого цвета. То есть развитие этих признаков определяется действием одного наследственного фактора (гена).
У человека встречается рецессивная наследственная болезнь — серповидно-клеточная анемия. Первичным дефектом этой болезни является замена одной из аминокислот в молекуле гемоглобина, что приводит к изменению формы эритроцитов. Одновременно с этим возникают нарушения в сердечно-сосудистой, нервной, пищеварительной, выделительной системах. Это приводит к тому, что гомозиготный по этому заболеванию ребенок погибает в детстве. Причиной синдрома Марфана является доминантная мутация гена, контролирующего одновременно рост, длину пальцев, формирование интеллекта и форму хрусталика. Для человека с этим синдромом характерен комплекс следующих признаков — высокий рост, очень длинные гибкие («паучьи») пальцы, повышенный интеллект, близорукость.
Плейотропия широко распространена. Изучение действия генов показало, что плейотропным эффектом, очевидно, обладают многие, если не все, гены.
Таким образом, выражение «ген определяет развитие признака» в значительной степени условно, так как действие гена зависит от других генов — от генотипической среды. На проявление действия генов влияют и условия окружающей внешней среды. Следовательно, генотип является системой взаимодействующих генов.
-
Перейти к лекции №19 «Генетика пола»
-
Перейти к лекции №21 «Изменчивость»
-
Смотреть оглавление (лекции №1-25)
Сцепление генов. Опыты Моргана
Сцепленные гены — любые гены, расположенные в одной хромосоме.
Группа сцепления — все гены, расположенные в одной хромосоме.
■ Количество групп сцепления равно числу пар хромосом (т.е. гаплоидному числу хромосом). У человека 46 хромосом, т.е. 23 группы сцепления.
■ Наследование признаков, за которые отвечают гены из одной группы сцепления, не подчиняется законам Менделя.
Опыты Т. Моргана (1911-1912 гг.): анализ наследования двух пар альтернативных признаков у мух-дрозофил — серого (В) и черного (b) цвета тела и нормальной (V) или укороченной (v) длины крыльев.
Первая серия опытов: скрещивание гомозиготной доминантной (BBVV) особи (с серым цветом тела и нормальной длиной крыльев) с гомозиготной рецессивной (bbvv) особью черного цвета с короткими крыльями. Все потомки F1, в соответствии с первым законом Менделя, являются доминантными гетерозиготными (BbVv) особями серого цвета с нормальными крыльями.
Вторая серия опытов: анализирующее скрещивание гибридов первого поколения — гомозиготной рецессивной (черной короткокрылой) самки (bbvv) с дигетерозиготным (серым с нормальными крыльями) самцом (BbVv). Если предположить, что два гена, относящиеся к разным аллельным парам, локализованы в разных хромосомах, то у дигетерозиготы следует ожидать образования (в равных количествах) четырех типов гамет: BV, bV, Bv и bv. Тогда, согласно третьему закону Менделя, в потомстве должны присутствовать четыре разных фенотипа в равном количестве (по 25%). В действительности присутствовали только два фенотипа (в соотношении 1 : 1).
■ Это означает, что доминантные гены В и V, относящиеся к разным аллельным парам, локализованы в одной хромосоме (из пары гомологичных хромосом) и попадают в одну гамету, а оба рецессивных гена в и v локализованы в другой хромосоме и вместе попадают в другую гамету. Поэтому у дигетерозиготного самца мухи-дрозофилы образуется не четыре типа гамет (когда гены расположены в разных хромосомах), а только два: BV (50%) и bv (50%), и, следовательно, потомки F2 будут иметь два сочетания признаков.
Третья серия опытов: проверка предположения о полном сцеплении генов путем анализирующего скрещивания дигетерозиготной (серой с нормальными крыльями) самки (BbVv) из поколения F1 с гомозиготным рецессивным (черным короткокрылым) самцом (bbvv) из родительского поколения. В результате были получены потомки четырех фенотипов в следующем соотношении: по 41,5% особей с серым телом и нормальными крыльями (генотип BbVv) и особей с черным телом и короткими крыльями (генотип bbvv), и по 8,5% серых короткокрылых особей (генотип Bbvv) и черных особей с нормальными крыльями (генотип bbVv).
Отсюда следует, что сцепленные гены, т.е. гены, локализованные в одной хромосоме, не всегда передаются вместе, т.е. сцепление может быть неполным. Это связано с явлением кроссинговера. вероятность которого в данном случае составляет 17%.
Множественный аллелизм
В природе широко распространено явление, когда один и тот же ген имеет не две, а более форм. У растений это проявляется разнообразием полосок на листьях и лепестках, у животных — различными сочетаниями окрасов. У людей ярким примером множественного аллелизма является наследование ребенком группы крови. Ее система обозначается АВО и контролируется одним геном. Его локус обозначают I, а аллельные гены — IA, IB, IO. Сочетания IO IO дают первую группу крови, IA IO и IA IA – вторую, IB IO и IB IB – третью, а IA IB – четвертую. Кроме того, у людей определяется резус. Положительный дают сочетания 2 аллельных генов с признаком «+» или 1+ и 1-. Отрицательный резус дают два аллельных гена с признаком «-«. Систему резус контролируют гены CDE, причем ген D часто вызывает резус-конфликт между плодом и мамой, если у нее кровь резус-отрицательная, а у плода резус-положительная. В таких случаях, чтобы успешно завершились вторая и последующие беременности, женщине проводят специальную терапию.
7.Множественный аллелизм. Генетика групп крови.
Множественный
аллелизм — различные состояния (три и
более) одного и того же локуса хромосом,
возникшие в результате мутаций.
Присутствие
в генофонде вида одновременно различных
аллелей гена называют множественным
аллелизмом. Примером этому служат разные
варианты окраски глаз у плодовой мухи:
белая, вишневая,
красная, абрикосовая,
эозиновая,— обусловленные различными
аллелями соответствующего гена. У
человека, как и у других представителей
органического мира, множественный
аллелизм свойствен многим генам. Так,
три аллеля гена I определяют групповую
принадлежность крови по системе АВ0
(IA, IB, I0). Два аллеля имеет ген, обусловливающий
резус-принадлежность. Более ста аллелей
насчитывают гены α- и β-полипептидов
гемоглобина.
Причиной
множественного аллелизма являются
случайные изменения структуры гена
(мутации), сохраняемые в процессе
естественного отбора в генофонде
популяции. Многообразие аллелей,
рекомбинирующихся при половом размножении,
определяет степень генотипического
разнообразия среди представителей
данного вида, что имеет большое
эволюционное значение, повышая
жизнеспособность популяций в меняющихся
условиях их существования. Кроме
эволюционного и экологического значения
аллельное состояние генов оказывает
большое влияние на функционирование
генетического материала. В диплоидных
соматических клетках эукариотических
организмов большинство генов представлено
двумя аллелями, которые совместно влияют
на формирование признаков. .
У
человека серией множественных
аллелейпредставлен ген, определяющий
группу крови. При этом гены, обусловливающие
группы крови А и В, не являются доминантными
по отношению друг к другу и оба доминантны
по отношению к гену, определяющему
группу крови О. Следует помнить, что в
генотипе диплоидных организмов могут
находиться только два гена из серии
аллелей. Остальные аллели данного гена
в разных сочетаниях входят в генотип
других особей данного вида. Таким
образом, множественный аллелизм
характеризует разнообразие генофонда
целого вида, т. е. является видовым, а не
индивидуальным признаком.
Система
групп крови АВО
Система
групп крови ABO — это основная система
групп крови, которая используется при
переливании крови у людей. Ассоциированные
анти-А и анти-В-антитела (иммуноглобулины),
обычно относятся к типу IgM, которые, как
правило, образуются в первые годы жизни
в процессе сенситизации к веществам,
которые находятся вокруг, в основном
таких, как продукты питания, бактерии
и вирусы.
Система групп крови ABO также присутствует
у некоторых животных, например, у обезьян
(шимпанзе, бонобо и горилл).
Наследование
групп крови системы АВО. В системе АВО
синтез агглютиногенов и агглютининов
определяется аллелями гена I:I0, IA, IB. Ген
I контролирует и образование антигенов,
и образование антител.
При этом наблюдается полное доминирование
аллелей IA и Ю над аллелем I0, но совместное
доминирование (кодоминирование) аллелейIА
и IB. Из-за кодоминирования наследование
групп крови системы АВО происходит
сложным образом. Например, если мать
гетерозиготна по II группе, крови (генотип
IAI0), а отец гетерозиготен по Ш группе
крови (генотип IBI0), то в их потомстве с
равной вероятностью может родиться
ребенок с любой группой крови. Если у
матери I группа крови (генотип I0I0), а у
отца IV группа крови (генотип IАIB), то в
их потомстве с равной вероятностью
может родиться ребенок или со П (генотип
IAI0), или с III (генотип IBI0) группой крови
(но не с I, и не с IV).


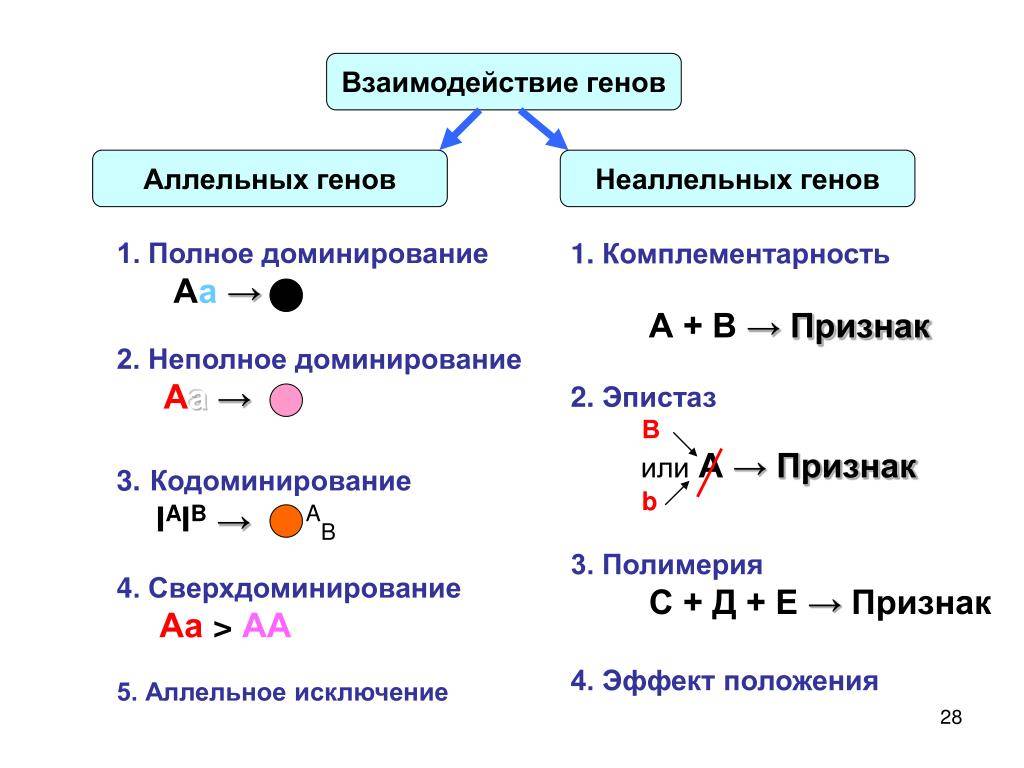
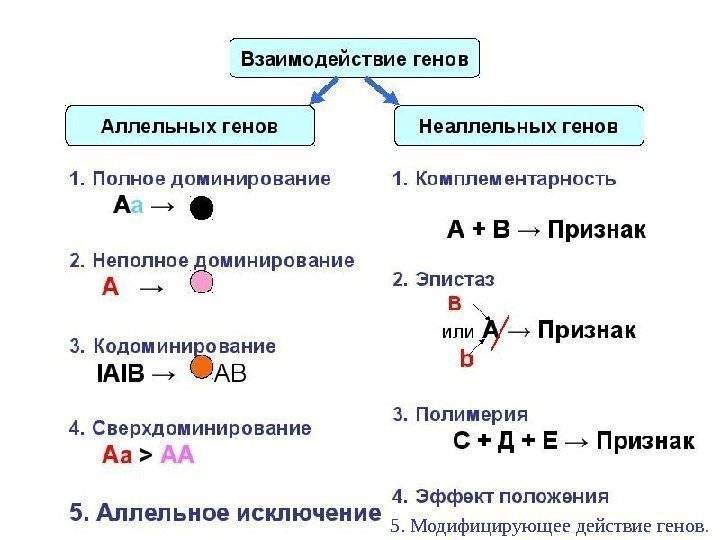
Виды
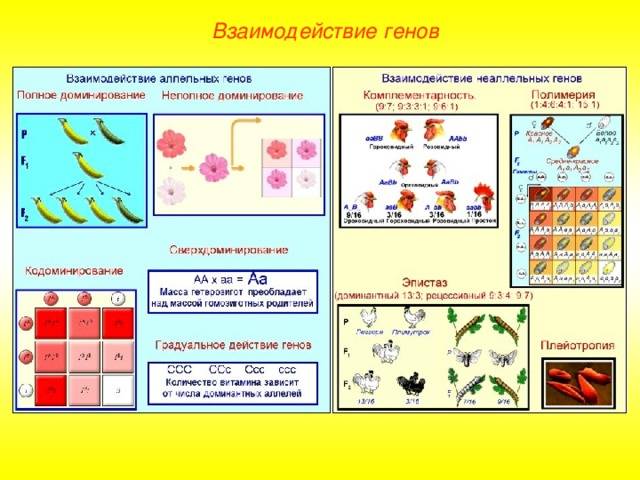
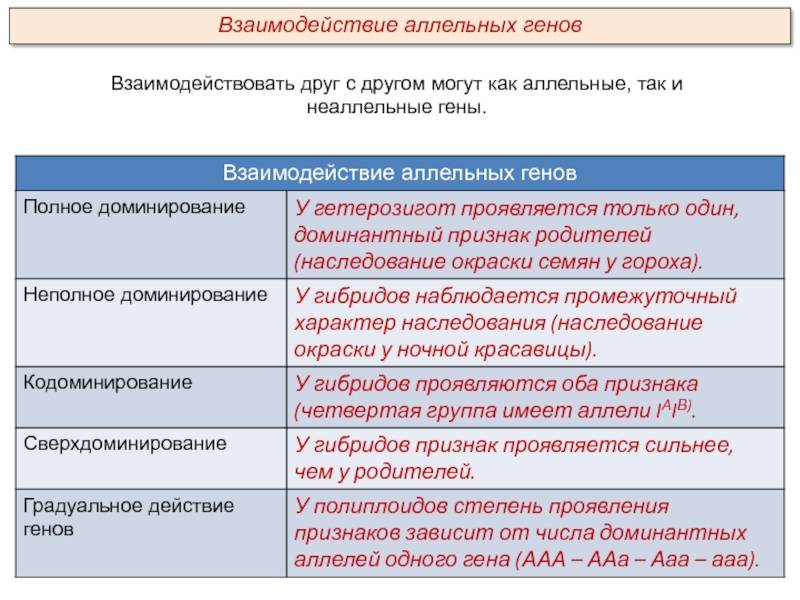
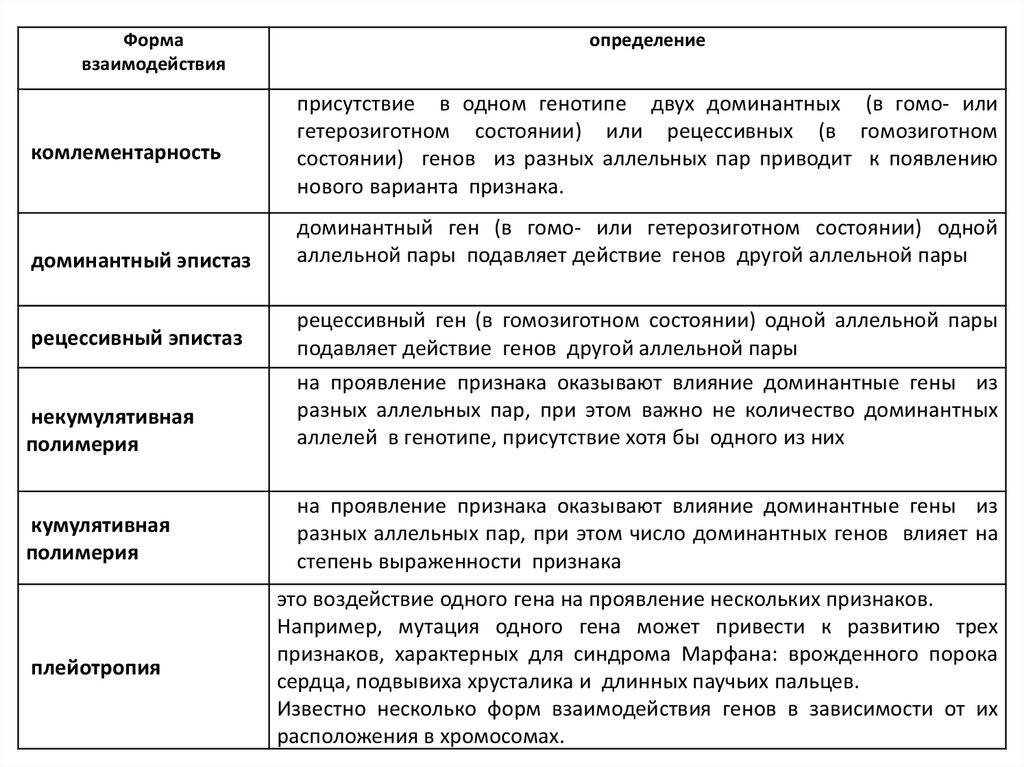
Основные типы взаимодействия аллельных генов представлены в таблице.
|
Тип |
Описание |
Пример |
|
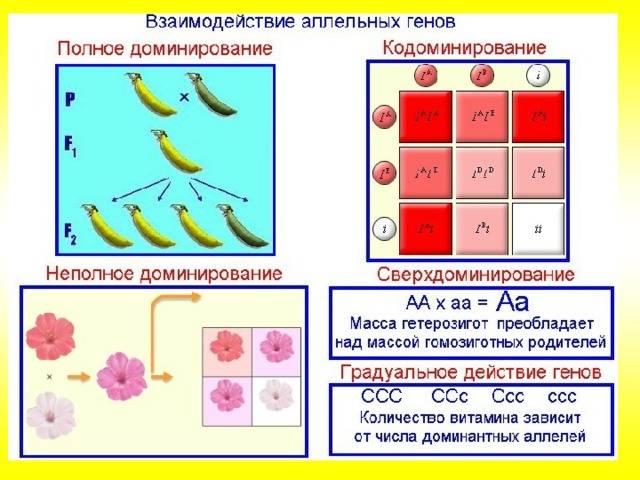
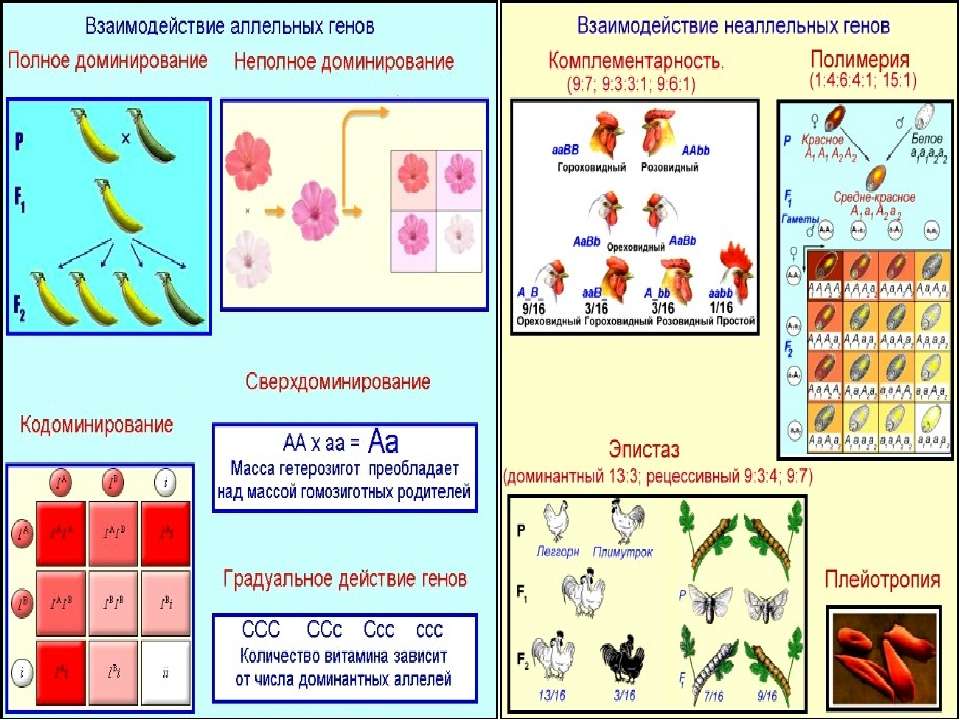
Полное доминирование |
Доминантный (преобладающий) ген полностью подавляет действие рецессивного гена при гомозиготном (АА) и гетерозиготном (Аа) наследовании. В фенотипе отражается только один признак, за который отвечает доминантный ген. Однако это не означает, что рецессивный признак полностью исключается. Расщепление признаков по фенотипу во втором поколении будет в соотношении 3:1, т.е. у одной особи полностью проявится рецессивный ген. Свойственно всем живым организмам, включая человека |
Положительный резус-фактор человека всегда доминирует над отрицательным, карий цвет глаз – над голубым, гладкая поверхность горошин доминирует над морщинистым |
|
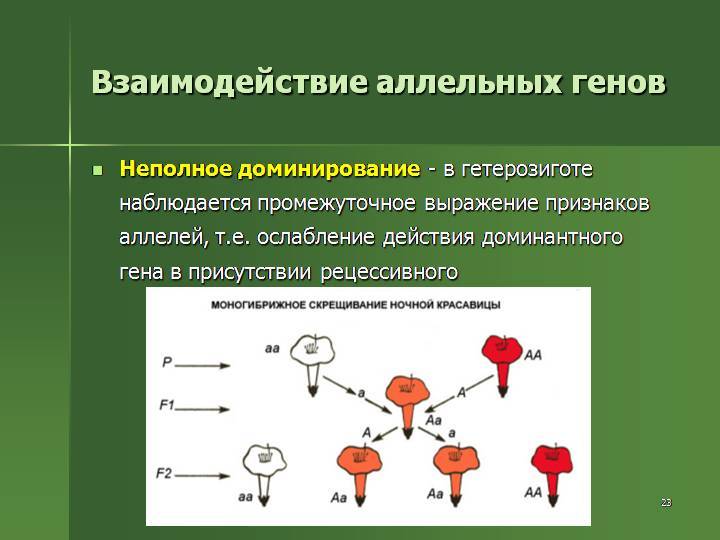
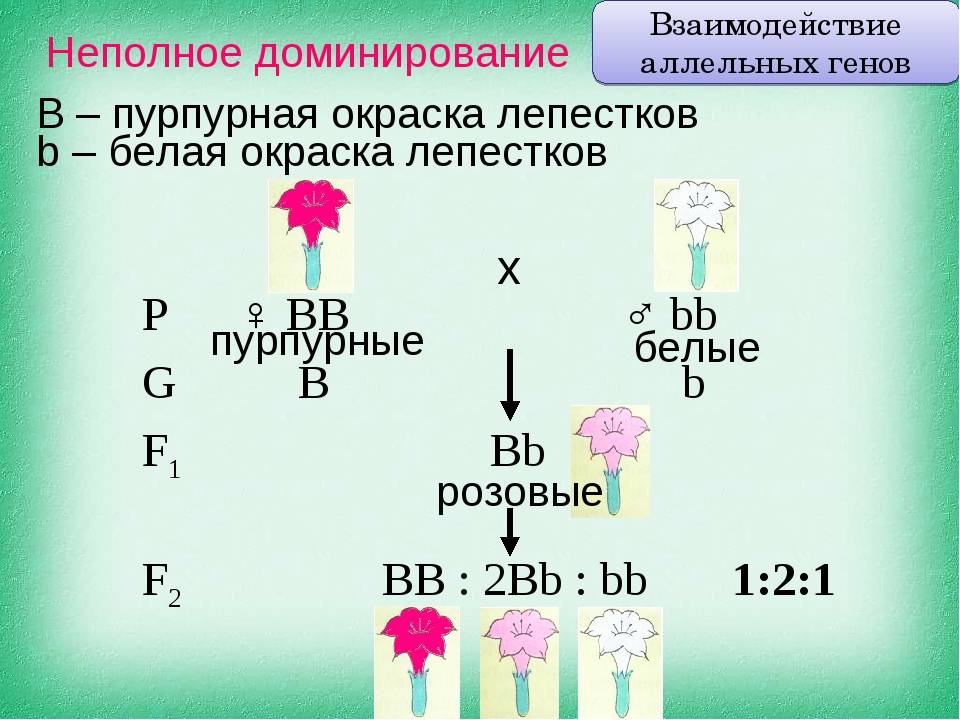
Неполное доминирование |
Промежуточное проявление признаков. Доминантный ген не полностью подавляет действие рецессивного гена. Это отражается на фенотипе в виде неполного проявления признака («усреднение» двух альтернативных признаков). Во втором поколении наблюдается расщепление по генотипу и фенотипу в соотношении 1:2:1 |
У ночной красавицы доминантный ген, отвечающий за красный цвет лепестков (АА), не полностью подавляет белый рецессивный ген (аа). В результате получаются гетерозиготные розовые цветки (Аа) |
|
Кодоминирование |
Полное проявление двух аллельных генов, в фенотипе у гетерозигот одновременно отражаются альтернативные формы признака, доставшиеся от родителей. В этом случае сложно определить доминантный и рецессивный гены, т.к. подавления признака не происходит |
Окраска шортгорнских коров: при скрещивании белых и красных коров получаются пятнистые особи с красно-белой окраской |
|
Множественный аллелизм |
Один и тот же ген может иметь в популяции более двух аллелей. Аллельные гены находятся в аналогичных участках хромосом. Множество аллелей обозначается индексами – А1, А2, А3 и т.д. Причина аллелизма – спонтанные изменения гена (мутации), сохраняемые в генофонде в ходе естественного отбора |
Дрозофила имеет 12 вариантов гена, отвечающего за цвет глаз, у человека три аллельных гена определяют группу крови |
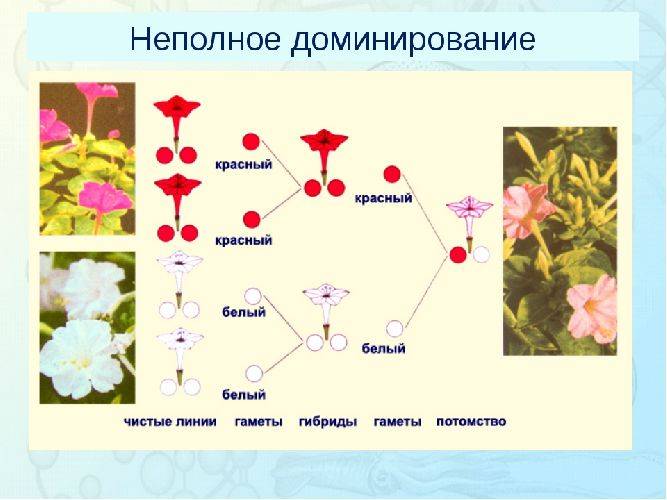
Рис. 2. Неполное доминирование.
Влияние каждого гена на признак зависит от ряда других генов. Поэтому современная генетика рассматривает генотип в целом, как целостную систему взаимодействия генов в комплексе.

Рис. 3. Шортгорнские пятнистые коровы.
Что мы узнали?
Рассмотрели виды взаимодействия аллельных генов. Выделяют полное и неполное доминирование, кодоминирование и множественный аллелизм. При полном доминировании доминантный ген всегда подавляет рецессивный. При неполном доминировании проявляется промежуточный признак. Кодоминирование предполагает одновременное проявление рецессивного и доминантного генов. Множественный аллелизм – вариации одного гена в генофонде популяции.
-
Вопрос 1 из 5
Начать тест(новая вкладка)
Экспрессия гена
Пенетрантность и экспрессивность. Норма реакции. Экспрессия генов — это процесс, в ходе которого наследственная информация от гена (последовательности нуклеотидов ДНК) преобразуется в функциональный продукт — РНК или белок. Экспрессия генов может регулироваться на всех стадиях процесса: и во время транскрипции, и во время трансляции, и на стадии посттрансляционных модификаций белков.






Регуляция экспрессии генов позволяет клеткам контролировать собственную структуру и функцию и является основой дифференцировки клеток, морфогенеза и адаптации. Экспрессия генов является субстратом для эволюционных изменений, так как контроль за временем, местом и количественными характеристиками экспрессии одного гена может иметь влияние на функции других генов в целом организме.
Экспрессивность и пенетрантность Под экспрессивностью понимают степень выраженности определенного признака.
Внешняя среда и гены-модификаторы могут изменить экспрессию гена, т. е. выражение признака. Изменчивость проявления мутантного гена у разных особей — довольно частое явление. Например, у потомства дрозофилы — мутантных «безглазых» мух с сильно редуцированным количеством фасеток — содержание их варьирует от почти полного отсутствия до половины нормы. Пенетрантность гена — это доля особей, у которых проявляется ожидаемый фенотип.
При полной пенетрантности (100 %) мутантный ген проявляет свое действие у каждой особи. При неполной пенетрантности (меньше 100 %) ген проявляется фенотипически не у всех особей. Экспрессивность и пенетрантность гена в значительной степени зависят, по-видимому, от влияния генов-модификаторов и условий развития особей.
Норма реакции — способность генотипа формировать в онтогенезе, в зависимости от условий среды, разные фенотипы.
Она характеризует долю участия среды в реализации признака и определяет модификационную изменчивость вида.
Виды
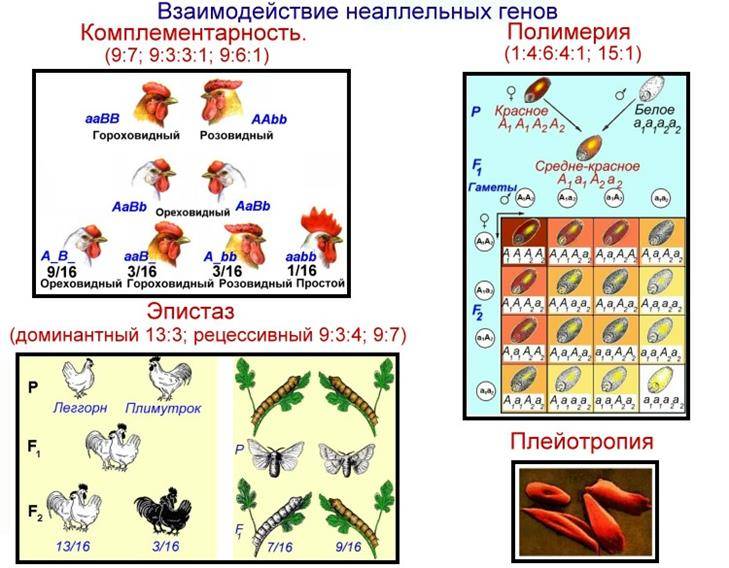
Существует несколько видов взаимодействия неаллельных генов, главные из которых подробно описаны в таблице.
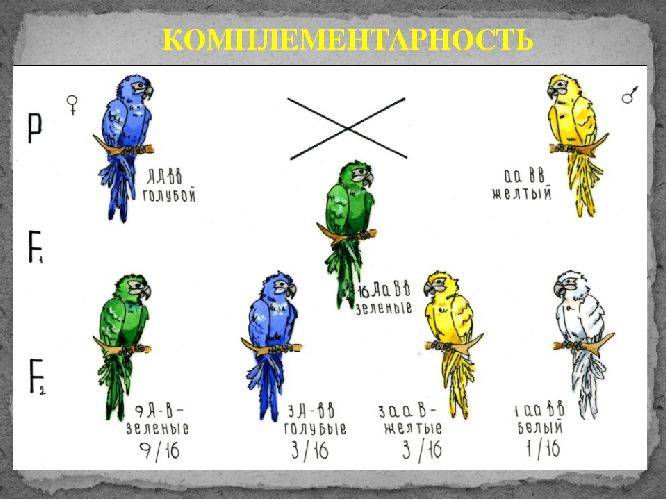
Рис. 2. Комплементарность.
|
Вид |
Описание |
Пример |
|
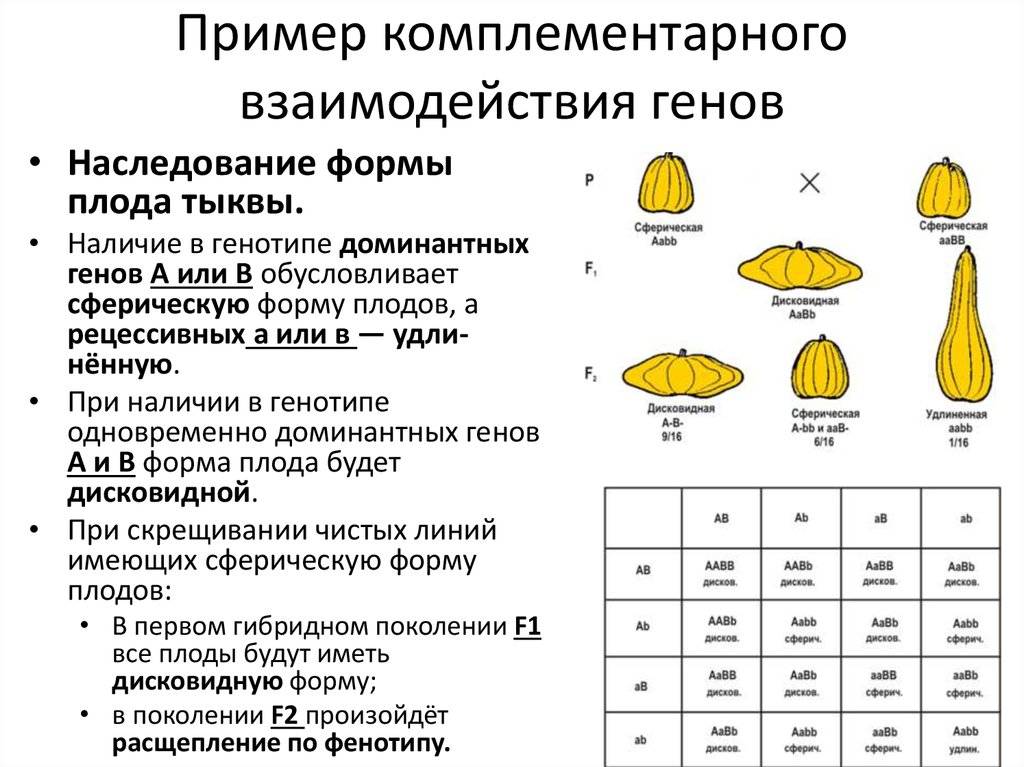
Комплементарность |
Признак, обусловленный двумя разными генами, проявляется только при сочетании двух доминантных аллелей. Такие гены называются комплементарными. Признак не формируется при отсутствии одного гена. Расщепление фенотипических признаков в F2 происходит в соотношении 9:7, 9:6:1, 9:3:4 |
Скрещивание душистого горошка с белыми цветками. В F1 все потомки имеют пурпурные цветки, т.к. сочетание доминантных генов А и В кодируют антоциан, придающий пурпурную окраску. По отдельности гены не образуют пурпур. В F2 происходит расщепление – 9 пурпурных (АВ), 7 белых (3 – Abb, 3 – aaB, 1 – aabb) |
|
Эпистаз |
Одна пара генов подавляет другую, не давая проявиться фенотипическому признаку. Подавляющий ген называется эпистатичным (ген-супрессор или ингибитор), подавляемый – гипостатичным. Ингибитор обозначается буквой I, i. Эпистаз может быть доминантным – подавление доминантным геном (I>B, b) и рецессивным – подавление рецессивным геном (i>B,b). При доминировании происходит расщепление генов в соотношении 7:6:3, 12:3:1, 13:3, при рецессивном проявлении – 9:3:4, 9:7, 13:3 |
Окраска овсяного зерна: А – чёрный цвет, В – серый цвет. В F1 все зёрна будут чёрного цвета, если ген А эпистатичен (AaBB или IiBB). В F2 произойдёт расщепление по цвету зерна – 12 чёрных, 3 серых и 1 белое. У 12 растений обязательно присутствует I-ген, у 3 он будет в рецессивном состоянии – i. Одному растению достанутся гены iibb (отсутствие чёрной и серой окраски), поэтому он будет белым |
|
Полимерия |
Количественные или мерные признаки, которые нельзя чётко разделить по фенотипу (рост, количество молока, жирность скота), определяются совокупностью генов. Выделяют кумулятивный и некумулятивный виды. В первом случае проявление признака зависит от суммы действий генов (чем больше доминантных генов, тем ярче признак). Во втором случае признак проявляется при доминантном гене, количество генов на проявление фенотипа не влияет. При кумулятивном виде в F2 наблюдается расщепление в соотношении 1:4:6:4:1, при некумулятивном – 15:1. Обозначают полимерные гены одной буквой (А, а, В, b и т.д.), а аллели – цифрой. Например, А1а1А2а2 |
Цвет кожи человека зависит от действия четырёх генов: А1А1А2А2 – чёрный цвет, а1а1а2а2 – белый, А1А1А2а2, А1а1А2А2, А1а1А2а2, А1А1а2а2, а1а1А2А2, А1а1а2а2, а1а1А2а2 – промежуточные значения от тёмного (почти чёрный) до светлого (почти белый) оттенка |

Рис. 3. Эпистаз.
Множественное действие генов называется плейотропией. Действие одного гена, как правило, обусловлено взаимодействием с другими генами. Таким эффектом обладает большинство генов, поэтому генотип представляет собой систему взаимодействующих генов.


Что мы узнали?
Узнали кратко о типах взаимодействия неаллельных генов. Существует три типа взаимодействия – комплементарность, эпистаз, полимерия. Для комплементарного проявления признака необходимо наличие двух доминантных генов. Для эпистаза характерно подавление одним геном действие второго гена. Полимерия – взаимодействие совокупности генов. Взаимодействие множества генов называется плейотропией.
Тест по теме
-
Вопрос 1 из 10
Начать тест(новая вкладка)
Комплементарность
При таком взаимодействии один доминантный ген дополняет другой, который также доминантный, но не является аллельным. Попадая вместе, они способствуют проявлению совершенно нового признака.
Можно привести пример проявления окраски у цветов душистого горошка. Наличие пигмента, а значит, окраски у цветка обеспечивается сочетанием двух генов — А и В. Если хоть один из них будет отсутствовать, то венчик будет белым.
У людей такое взаимодействие неаллельных генов наблюдается при формировании органа слуха. Нормальный слух может быть только, если присутствуют оба гена — D и E — в доминантном состоянии. При наличии только одного доминантного или обоих в рецессивном состоянии слух отсутствует.
Взаимодействие генов в детерминации признаков. Виды взаимодействия генов.
Но в диплоидном организме может быть только 2 члена из серии.
У человека с серией множественных аллелей связано наследование групп крови. Ген 1А — отвечает за выработку в эритроцитах агглютиногена А.
Ген 1В — отвечает за выработку в эритроцитах агглютиногена В.
Ген I — ни А ни В не вырабатывается.
I (О)- генотип I0 I0
II (A)- IAIA; IAI0
III (В)- I B I B; I B I0
IV (АВ)- I A I B;
Наследование групп крови нужно знать для определения вероятной группы крови у потомков, зная группы крови родителей; для правильного переливания крови.
Взаимодействие неаллельных генов.
Эпистаз — это такое взаимодействие между неаллельными генами, когда один неаллельный ген подавляет действие другого неаллельного гена. Ген, который подавляет называется эпистатический,а ген, который подавляется — гипостатический.
Различают доминантный эпистаз: доминантный аллель одного гена подавляет действие и доминантного, и рецессивного аллеля другого гена.
Например: наследование окраски у плодов тыквы.
А — ген, отвечающий за жёлтую окраску,
а — ген, отвечающий за зелёную окраску.
При генотипе Аа и АА тыквы жёлтого цвета, при генотипе аа- зелёного.
Если в генотипе растения появляется особый доминантный (эпистатический) аллель другого гена В, в гомо- или в гетерозиготном состоянии, то он подавляет действие «А» и «а» и развивается белая окраска плодов тыкв.
Генотипы растений с белыми плодами:
ААВВ, АаВВ, ААВв, АаВв, ааВВ, ааВв
Рецессивный эпистаз: рецессивный аллель эпистатического гена, если он в гомозиготном состоянии, подавляет действие доминантного и рецессивного аллеля гипостатического гена.
Например: у человека с генотипом I BI0, которому соответствует III (В) группа крови, фенотипически проявляется I (О) группа.
Это происходит потому, что в генотипе есть рецессивный эпистатический ген в гомозиготном состоянии, который подавляет действие гена I B .
Комплементарность — это такое взаимодействие между неаллельными генами, когда признак проявляется только при одновременном присутствии в генотипе двух доминантных аллелей неаллельных генов.
А-В- при таком генотипе признак проявляться будет.
ааВ- Признак проявляться не будет.
аавв
Например: наследование серой окраски у мышей происходит, если в генотипе одновременно присутствуетдоминантный аллель гена, отвечающего за синтез пигмента (А), и доминантный аллель гена, отвечающего за распределение этого пигмента у основания волоса и на его конце (В).
Полимерия— это явление, когда один признак определяется несколькими неаллельными генами.
Полимерия регулируетразвитие количественных признаков. Например, у человека интенсивность пигментации кожи определяется 4 неаллельными генами.



При генотипе A1A1A2A2A3A3A4A4 цвет кожи очень тёмный, при генотипе — а1а1а2а2а3а3а4а4 светлый, а при генотипе A1а1A2 а 2A3 а3A4а4 — средняя интенсивность окраски кожи.
Экспрессивность — качественный показатель фенотипического проявления гена.
Пенетрантность — количественный показатель фенотипического проявления гена.
Он выражается в % и говорит о том, у какого количества особей этот признак проявляется фенотипически.
1 ген может отвечать за развитие 1 признака — моногенность.Если 1 ген отвечает за развитие нескольких признаков, то это плейотропия,т.е. множественное действие гена.
Например: у человека 1 доминантный мутантный ген отвечает за развитие синдрома Марфана: длинные пальцы, подвывих хрусталика глаза, расширение аорты.